Essays on Dog Genetic Diversity
Inspired by conversations hosted by Jemima Harrison
The purpose of this website is to host essays and information about maintaining viable populations of healthy pure bred dogs
The purpose of this website is to host essays and information about maintaining viable populations of healthy pure bred dogs
Each cell within the body is composed of cytoplasm, a jelly-like layer of material that surrounds a nucleus. Within the nucleus are a number of threadlike chromosomes that are almost entirely made up of two kinds of chemical substances: nucleic acids and proteins. Nucleic acids have at least two functions: to pass on hereditary characteristics and to trigger the manufacturing of specific proteins. The two classes of nucleic acids are the deoxyribonucleic acids (DNA) and the ribonucleic acids (RNA). DNA, the genetic building block, is made up of substances called nucleotides, each of which consists of a phosphate, a sugar known as deoxyribose and any one of four nitrogen-containing bases. These four nitrogenous bases are adenine (A), thymine (T), cytosine (C) and guanine (G). Canine DNA is about 6 billion nucleotide pairs long. Each base is attached to a sugar molecule that is linked by a hydrogen bond to a complementary base on the opposite strand. These bases are complementary because only adenine pairs up with thymine, and only cytosine pairs up with guanine; thus the pairs are AT and CG.
In all mammals, the DNA molecule appears as two complementary strands that are wrapped around each other like the railings of a spiral ladder, known more formally as the double helix of Crick and Watson. 4 The strands (sides of the ladder) are composed of alternating phosphate and sugar molecules. The nitrogen bases, joining in pairs, serve as the rungs. The two strands are held together by weak electrical bonds between the bases on each strand/rung, thus forming base-pairs. Each strand has its own polarity opposite of the other. Thus if you turned the strands upside down, the picture would not change. An easy way to visualize the opposite polarity aspect of the two chains is to think of two identical snakes intertwined around each other but facing opposite directions (head to tail and tail to head). Thus, each half of the double helix serves as a genetic template of its complementary half.\\
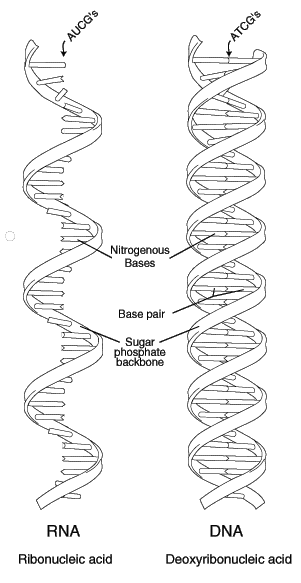
Before a cell can express a particular gene, it must first transcribe that specific part of the DNA into messenger ribonucleic acid (mRNA). This is similar to the formation of a complementary strand of DNA during cellular division, except that RNA contains uracil (U) instead of thymine as one of its four nucleotide bases. In the process of transcribing DNA into mRNA, all the T bases are converted to U bases. These bases C, G, A, and U are the alphabet of the genetic code. A sequence of AGT in the coding strand of the DNA thus produces a sequence UCA in the mRNA.
Think of codons as three-letter “words” identifying the bases DNA uses to specify particular amino acids as building blocks of proteins. Normally, codons signal the initiation of a protein chain, its end or a particular amino acid. For example, CUU stands for the amino acid leucine. CUA, CUG, and CUC also “code” for leucine, so there is some redundancy in the system. Notice in this example that it is only the last base that is different (U vs. A vs. G vs. C). The term degeneracy is used when a change in a base does not affect the amino acid being added to the polypeptide.
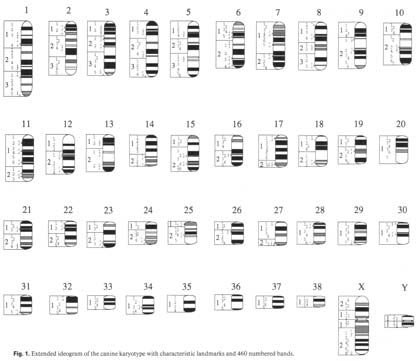
What is a gene? A gene is the basic unit of inheritance. Genes contain a set of directions for producing a bit of RNA, a protein or a polypeptide. If all goes well, a complete set of genes - one half from each parent - is inherited. If the two copies of each gene are exactly alike, the progeny are homozygous at that locus. When homozygous, only one form of the gene will be passed on. If the gene inherited from one parent is different from the gene inherited from the other, the progeny are heterozygous. In this case, there is an equal chance that one or the other form of the gene will be passed on. Different forms of the same gene are called alleles. In the dog, the various genes are located among 78 different chromosomes. What is not known is how many genes exist, although a rough estimate has been made that there may be as many as 100,000 or as few as 30,000. It is also not known where on the various chromosomes specific genes are located. In fact, scientists have just recently karyotyped the canine. This provides the ability to differentiate between specific chromosomes. This will be valuable information when scientists are finally able to map the canine chromosome. Such a genetic map will not only allow the determination of the position of genes relative to each other, but also will reveal their approximate distance from each other. Mitosis is the process of one cell splitting and becoming two cells. This act of division is the result of a series of events known as the “cell cycle,” which consists of several distinct phases or stages. The resting or quiescent state between cellular divisions is called the “G0” stage. “G1” is the phase in which all the cellular proteins needed for mitosis are made and is the first control point where the cell must “decide” to move on to the next stage--the “S” phase. During the “S” phase, the cell’s genetic material is duplicated so each of the daughter cells is genetically identical to the parent cells--unless something goes wrong. The S phase is so named because this is the point during which new DNA is synthesized. The period from the end of the S phase until the actual division of the cell is known as the “G2” stage and is the second control point during which a decision is made whether or not to actually undergo division. Mitosis is further divided into prophase, metaphase, anaphase, and telophase. The stages G1, S and G2 constitute the interphase. In a process called meiosis, germline cells--sperm and eggs- go through one more division in which their DNA is not duplicated. This leaves them with only half the normal number of chromosomes that somatic cells have. Somatic cells are all the cells of the body except the germline cells. Once these two cells combine to form a fertilized egg or zygote, they then have the proper amount of DNA--half from their mother and half from their father. This is important because mutations in the germline cells are passed on to the offspring, whereas those that occur in somatic cells only affect the individual.\\ BREAKING DOWN THE GENE
THE CELL CYCLE
2-2 Stages of the cell cycle ====

Mutations (changes in the genes) are caused by a variety of mechanisms. Some of the most common are mistakes made when the organism’s DNA is replicated prior to a cell dividing. Although there are body system safeguards in place to prevent this from happening, nothing is foolproof, and eventually over time, failure to replicate DNA accurately will occur. Likewise, errors can occur all along the pathway that leads to the translation of messenger RNA into a specific protein. These errors can occur spontaneously or be the result of exposure to natural and/or man-made mutagens. Certain chemicals or exposure
to certain types of radiation can cause genetic changes. What is important to remember is that these mutations are random events with respect to their adaptive potential. In other words, they will happen independently of whether they have beneficial or harmful consequences. More often than not these mutations are harmful, as they are changes to the make up of a living organism. Just how harmful depends upon the type of mutation that occurs and the environment in which they occur. Most mutations fail to thrive, reproduce or survive and thus are not passed on to successive generations. There are several kinds of gene mutations, each having a unique range of potential effects. This is important to recognize because many genetically transmitted diseases result from a specific kind of mutation. Each of these forms of mutation is the result of the organism failing to reproduce its DNA accurately all of the time and subsequently passing these genetic changes to successive generations. The result of this type of mutation can range from no effect at all to one that has severe consequences to the affected organism. Remember DNA is made up of four different nucleic acids: thymine (T), adenine (A), guanine (G) and cytosine (C), and remember that
thymine always pairs up with adenine and guanine always pairs up with cytosine. Hence the name base-pair mutation. Sometimes when the DNA strand is being replicated the wrong base is inserted. This can result in a different amino acid being added to the protein being made. If the essential biological function of that protein is not changed then there is no detectable effect. However, if the substitution affects the active site of an important enzyme or changes it’s three-dimensional shape, then it modifies the fundamental nature of the protein. If this occurs along an essential metabolic pathway, the results can be disastrous. The most unfortunate result of a base-pair substitution is when this mutation codes for a stop codon. Keep in mind that a codon is that portion of the messenger RNA that codes for a specific amino acid. A start codon serves rather like a capital letter indicating the start of a sentence. A stop codon is one that does not specify an amino acid, and serves much as a comma or a period punctuating the genetic message. If, by chance, a mutation produces one of the stop codons, than the process of making the protein is terminated.
An example of this type of mutation is the one that leads to a form of progressive retinal
atrophy (PRA) in the Irish Setter. A substitution of an A for a G produces the stop codon
(TAG) that replaces the normal codon for the amino acid tryptophan (TGG). This
prevents a protein called PDEB (phosphodiesterase beta) from being produced in its full- length form. The shortened protein is unstable and is degraded within the retinal cells in which it is needed. The lack of this protein causes the retina to degenerate, resulting in blindness in those Irish Setters that have two copies of the mutant gene, and no normal copy. In the normal cell replication process, DNA is transcribed into messenger RNA, which in turn is translated into a series of amino acids. This always occurs in a specific manner,
i.e., it always begins at a definite spot and it is ‘read’ in multiples of three and in a particular orientation along the length of the strand of DNA. This is called a reading frame. If there is an addition or deletion of one or two base pairs, then the result is often a very altered sequence of amino acids in the final protein product resulting in what is termed a frameshift mutation. An example of this is the mutation that leads to an inherited form of anemia in Basenjis. A deletion of a single nucleotide in the 433rd codon of the gene encoding a protein called PK (pyruvate kinase) causes a shift in the reading frame. The misformed and shortened protein (a new stop codon is ultimately encountered) is unstable in the red blood cells that carry oxygen throughout the body. The lack of the PK causes the red blood cells to slowly be destroyed and results in the anemia. Molecular geneticists used to think that all of the DNA coding for a particular protein was continuous until they started to look at more complex organisms. What they found, in these types of cells, is that the DNA that makes up a gene is often distributed in discontinuous sections called exons, interspersed with long segments of non-coding DNA known as introns. These sections are transcribed into messenger RNA along with the exons, but before the RNA is translated into a protein they are ‘edited’ or ‘spliced’ out. A change of even a single nucleotide in one of the exons of the gene can cause a shift or alteration of the splice-site. A genetic disease that affects Dobermans is a perfect illustration of this type of mutation. von Willebrand disease is a bleeding disorder that effects the animal’s ability to form blood clots. Other breeds also have this disease, but what had perplexed those doing von Willebrand research was that Dobermans appeared to have a milder form of the disease. The discovery of a splice-site mutation that codes for von Willebrand factor has cleared up their mystery. George Brewer MD of the University of Michigan suggests that one use the following analogy in order to explain how the mutation functions5. Imagine that a freight train is supposed to go from point A to point B along a railroad track. Somewhere between A and B is a spot where a sidetrack goes to point C. Normally, the train never goes to point C because the switch, that connects the two tracks, is never thrown. If the switch is broken (the mutation) then the lock that prevents the track from connecting to point C is no longer effective. The switch can now toggle back and forth, sending some trains to point B and some trains to point C. In affected Dobermans, the defective switch sends the train to the wrong destination about 90-95% of the time, the train rumbles over the cliff and is never heard from again (i.e., the proper protein is never made). However, sometimes the switch jiggles the right way and the train ends up at the normal destination and the proper protein is made. If both copies of the gene are mutated, then each gene can make the right protein about 5 to 10% of the time. Affected Dobermans are thus producing von Willebrand factor at least some of the time and so their symptoms are not as severe. A mystery explained. Other types of mutation occur during cellular division because of chromosomal abnormalities. Let’s review how chromosomes are normally duplicated during cellular division. Keep in mind that the prophase is the first stage of cell division. The nucleus swells and the chromosome becomes visible. During interphase the DNA has been duplicated and consists of two linked (sister) chromatids held together at a centromere. A structure called a centriole appears and moves towards the opposite poles of the nucleus. The next stage is called the metaphase. During this period the spindle fibers are formed and are attached at the centromeres and the chromosomes line up along the equator. This is the very best time to examine the complete set of chromosomes within a cell. If a cell is “fixed” at this point in the cycle and stained with special dyes, a cytogeneticist can determine if there is the correct amount of DNA, any deletions or other abnormalities and the sex of the individual. This “picture in time” is called a karyotype.
Following the metaphase is the anaphase. During this stage, the sister chromatids separate and migrate to opposite ends of the cell, the nuclear membrane has disappeared and the cell elongates as the diameter of the cell decreases at the equator. At telophase a new membrane is formed about the two new cells, the chromatids (now called chromosomes) uncoil and the nucleus is reformed. In sum, chromatids are just compacted chromosomes. Before looking at the different types of chromosomal abnormalities let us first discuss some of the terms used to describe them. Chromosomes are either sex chromosomes, in mammals these are the well-known X or Y chromosome, or autosomes. Autosomes are any chromosome that is not a sex chromosome. Chromosomes can be divided further into metacentric, submetacentric and acrocentric. These terms describe the position of the centromere within the chromosome. Metacentric chromosomes have their centromere near the center of the chromosome. Those chromosomes whose centromeres are slightly off-center are referred to as submetacentric, while acrocentric chromosomes have their centromere located close to one end. Dog chromosomes are mostly acrocentric. The small “arm” of the chromosome is referred to as p for “petit” and the longer arm is called q – the next letter in the alphabet. Another naming protocol is to designate regions and bands from the centromere outward. Depending on which staining techniques are used, banding patterns are seen that are characteristic for each chromosome pair. Thus, the designation 7q31.2 refers to the long arm of chromosome 7, region 3, band 1, sub-band 2. Normally dogs have 39 chromosomes in their germline cells, i.e., sperm and eggs. Germline chromosomes are haploid, i.e. they contain one copy of each chromosome.Somatic cells are diploid – they contain two homologous copies of each chromosome. A failure of the chromosome to separate properly during cell division (nondisjunction) can lead to a decrease or increase in the number of normal chromosomes. For example: Triploidy is the presence of three haploid sets of chromosomes, instead of two. Monosomy is the situation where a diploid cell, in which normally one or more of the homologous chromosome pairs is represented, only has one chromosome of the pair. Trisomy is the term used which indicates the presence of an extra whole chromosome. Each canine somatic cell usually has 78 chromosomes (2x39), but in trisomy, this is increased to 79. Down’s syndrome in humans is caused by this anomaly as there is an extra chromosome number 21. A chromosomal deletion occurs when part of a chromosome is missing. The damage that can occur depends upon how big of a piece is missing and where the deletion occurs. A chromosomal duplication happens when a section of the chromosome is reproduced twice. Depending on what section is duplicated there can be extra sets of genes present that can cause birth defects or developmental problems. A chromosomal ring occurs when the q and the p ends stick together. This can cause loss of information and/or cause problems when the cell divides. A chromosomal inversion is caused when there are two breaks in one chromosome and the area between the breaks are turned around and reattached. If the break includes the centromere, it is called a pericentric inversion. If not, it is a paracentric inversion.
Chromosomal translocations are rearrangement of a chromosome in which a segment is moved from one location to another, either within the same chromosome or to another chromosome can be balanced or unbalanced. A balanced translocation occurs when the same piece of chromosome, say the q arms of two different chromosomes, are broken off and attached to the other chromosome. No genetic material is lost, it is just on a different chromosome. This should not cause problems with the individual but could with that individual’s progeny. An unbalanced translocation, occurring in a germline cell, results in 3 copies of a section of chromosome in one cell and only one copy in the other. Both trisomy and translocation are implicated in canine cancers. The other chromosomal anomalies are well characterized in human disease but not in the canine. Chapter01[Previous Section]||
Chapter03[Next Section]GENE MUTATIONS
Base-Pair Substitution Mutations
Frameshift Mutations
Splice-Site Mutations
CHROMOSOMAL ABNORMALITIES
2-3 Canine Karyotype
Numerical chormosomal abnormalities
Structural anomalies
Pedigree Dogs Exposed - The Blog
A Bitter Cynoanarchist Rages On
Online Mendelian Inheritance in Animals (OMIA) OMIA
Understanding Gene Loss in successive generations An Interactive Academic Site on "Gene Dropping" at University of Montana
GENUPComputer aided learning for quantitative genetics.
Although a Windows application Bonnie Dalzell has been able to run it under Ubuntu Linux from my desktop.
A tactical approach to the design of crossbreeding programs.
Cultivar Crazy - Preserving the Genetic Heritage of Plants
Applied Animal and Plant Breeding
The Effects of Genetic Drift, Inbreeding, and Interpopulation Hybridization experimental study involving Drosophila (fruit flies)
Inherited Diseases in Dogs (IDID)
Canine Inherited Disorders Database
List of Inherited Disorders in Animals (LIDA)
VMDB Veterinary Medical Database
The Veterinary Medical Database (VMDB) compiles patient encounter data from nearly all North American veterinary medical databases.
CERFCanine Eye Registration Foundation
CHIC) Canine Health Information Center
OFAOrthopedic Foundation for Animals
CERF Canine Eye Registration Foundation
A Healthier Future for Pedigree Dogs
Closed Registries, Genetics, and Inbreeding Depression
A series of articles concerning the genetics of selection, inbreeding, inheritance of congenital defects.
All 4 essays by Susan Thorpe-Vargas Ph.D., John Cargill MA, MBA, MS, and D. Caroline Coile, Ph.D
Project to breed Dalmations with normal uric acid metabolism
Clarmorris Parson Russell Terriers
The Effects of Genetic Drift, Inbreeding, and Interpopulation Hybridization experimental study involving Drosophila (fruit flies)
« Back to DogDimension.org ] [ Hosting courtesy of Boarding at the Wedge
The layout for these essays is based on a css template by Matthew James Taylor, available via this link at: MatthewJamesTaylor.com although I [Bonnie Dalzell] have modified the template to some extent.